
Intensiv träning aktiverar proteinsyntes, men bara om rätt näring finns för att stödja den. Om du läser artikeln John Meadows och jag skrev som heter Maximize Protein Synthesis, eller om du har läst något som T Nation har sagt om ämnet i flera år, är du troligen bekant med begreppet ”det anabola fönstret” och vikten peri-träning näring.
Vad som händer på mobilnivå timmarna efter träning är förutsägbart för långsiktiga vinster. Utnyttja detta "anabola fönster" så växer du som aldrig förr. Missa det konsekvent, och, lycka till.
Att få rätt makronäringsämnen vid rätt tidpunkter är nyckeln, men makron är bara en del av helheten. En viktig men ändå ofta förbises aspekt av muskelproteinsyntes är cellvolym. Cellvolymen är inte bara kosmetisk; det är den främsta drivkraften för aminosyratransport, och arbetar också bakom kulisserna för att aktivera proteinsyntes och undertrycka proteinnedbrytning.
En full / volymiserad muskel är en anabol muskel. Medan vi har vetat i över 20 år att cellsvullnad hämmar proteinnedbrytning och stimulerar proteinsyntes i vissa celler (1-3), var mekanismen som länkar cellvolym till proteinsyntes fram till nyligen ett mysterium.
Vad vi vet nu är att proteinsyntes styrs av enzymet mTOR, som aktiveras av mekanisk stress, tillväxtfaktorer och leucin.
Medan alla dessa tre är viktiga för träningsstimulans, är mTOR-signalering också beroende av cellvolym.(4) Detta är särskilt viktigt i skelettmuskler, där cellvolymisering aktiverar glykogen- och proteinsyntes och hämmar proteinnedbrytning.(5, 6)
Det vetenskapliga genombrottet som ledde till sambandet mellan cellvolym och proteinsyntes inträffade 2005, när en grupp forskare upptäckte att det krävs mer än bara leucin för att aktivera mTOR - glutamin krävs också.(7)
Detta kom som en överraskning. Även om glutamin anses vara en "villkorligt essentiell" aminosyra som begränsar proteinnedbrytningen under svårt trauma eller stress, hade det aldrig kopplats till mTOR-aktivering.
Glutamin var nödvändigt för leucinupptag och cellvolymisering, vilka båda behövs för att aktivera proteinsyntes. Författarna fortsatte med att visa den cellulära glutamin tömning resulterar inte bara i en minskning av cellvolymen, utan minskar också leucins förmåga att aktivera proteinsyntes.(7)
Denna upptäckt var enorm, eftersom den gav en direkt koppling mellan glutamin, cellvolymisering och proteinsyntes. För första gången visades att glutamin är nödvändigt för aktivering av proteinsyntes med leucin.
Medan denna studie föreslog att glutamin är en mycket viktig bit i pusslet som kopplar samman cellvolym och proteinsyntes, fungerade den exakta mekanismen inte förrän 2009, då Nicklin et al. upptäckte att glutaminexport är kopplat till leucinimport och mTOR-aktivering.(8)
För att få leucin in i cellen finns det en första period med ”glutaminbelastning.”Detta drar också in vatten, vilket ökar cellvolymen. Efter "glutaminbelastning" -fasen exporteras glutamin ut ur cellen i utbyte mot import av leucin.
Nicklin et al. upptäckte också att cellulära glutaminnivåer är hastighetsbegränsande för aktivering av proteinsyntes med leucin. När celler samtidigt behandlades med glutamin och en EAA-blandning innehållande leucin försenades aktivering av proteinsyntes i 60 minuter. När samma celler glutamin "förladdades" aktiverades proteinsyntes inom 1-2 minuter efter att cellerna fick leucin.
Detta resultat var viktigt eftersom det förklarade fördröjningstiden för aktivering av proteinsyntes med leucin i denna experimentella modell.
Dessa resultat belyser äntligen det cellulära maskineriet som reglerar aminosyratransport och hur det kopplas till kontrollen av proteinsyntes.
Detta arbete måste dock tolkas med en viss försiktighet. En viktig varning för dessa studier var att de utfördes in vitro (i.e. i cellkulturer) där reglering eller proteinsyntes är mycket enklare. Muskelceller kan göra glutamin efter behov från andra aminosyror, och "glutaminutarmning" i denna cellodlingsmodell är inte representativ för mer fysiologiska situationer in vivo.
Höga proteinsyntes kan inte upprätthållas på obestämd tid i muskelvävnad med eller utan glutamintillskott. Glutamin kan dock användas för att strategiskt stödja proteinsyntes genom att optimera cellvolym under post-workout-perioden.

Celler är mycket upptagna, och det finns många membranbundna jonkanaler och transportproteiner som reglerar trafik in och ut ur cellen. Det finns särskilt två klasser av aminosyratransportörer som är viktiga här: "System L" och "System A" aminosyratransportörer är närmast kopplade till mTOR-signalering och proteinsyntes.(8-10)
Aktiviteten hos System A- och System L-transportörer är kopplade, vilket gör att leucin och andra BCAA kan absorberas i cellen.(11) System L-transportörer ansvarar för tillströmningen av leucin och andra BCAA i utbyte mot utflödet av glutamin.
System A-transportörer arbetar dock genom en annan mekanism där glutamin är kopplat till natriumupptag.(12, 13) Kopplingen mellan natriumupptag och System L / System A-aminosyratransportörer kallas Tertiär aktiv transport (kortfattat TAT). Det är TAT som i slutändan driver leucin inuti cellen vilket leder till mTOR-aktivering och proteinsyntes.(11)
Du kan se hur TAT fungerar i figuren nedan:
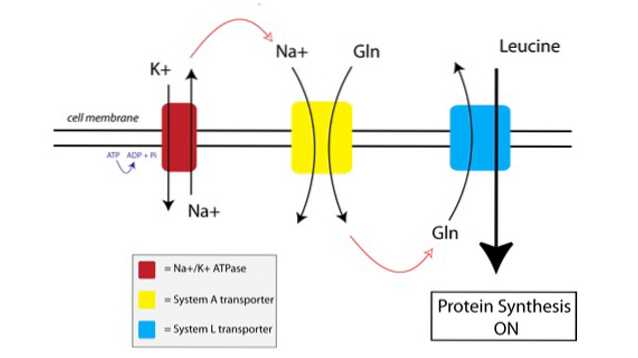
För det första använder en membranbunden pump som kallas natrium-kalium ATPas-pump (Na + / K + ATPas, röd i figuren ovan) energi från ATP för att flytta natrium utanför cellen, mot dess koncentrationsgradient.
Den ökade koncentrationen av natrium utanför cellen kopplas till importen av glutamin av System A-transportören (gul i figuren). Glutamin och natriuminflöde i cellen drar också in extra vatten, vilket får cellen att svälla. Detta sätter cellen i ett anabolt tillstånd, vilket grundar proteinsyntesmaskineriet för aktivering.
När glutamin byggs upp till tillräckligt höga nivåer inuti cellen aktiveras System L-transportörerna (blå i figuren), som skyttlar glutamin utanför cellen i utbyte mot leucintag. Inträde av leucin i cellen är utlösaren för proteinsyntes.
Även om detta hittills har varit en bra lektion i biokemi är upptäckten av TAT inte bara viktigt för cellbiologer. Nu när vi vet hur cellvolymen är kopplad till aminosyratransport och proteinsyntes kan vi utforma flera näringsstrategier för att maximera processen när den räknas - under den kritiska perioden efter träning.
Proteinsyntes är helt beroende av cellhydrering - om du ens är lite uttorkad är möjligheten att återhämta sig efter intensiv träning helt komprometterad. Att få massor av vatten är ingen idé här, men bara vatten räcker inte.
Elektrolyter som natrium, kalium, klorid och fosfat fungerar också som ”osmolyter” eftersom de drar vatten in i cellen. Efter en intensiv träningspass behöver vi vatten, aminosyror och elektrolyter för att maximera cellvolymiseringsprocessen som driver proteinsyntes.
Natrium, magnesium, kalcium, kalium, fosfat och klorid (för att nämna några) är alla viktiga här. På en minimal nivå (såvida du inte gör det enligt råd från din läkare), var inte undan natrium före eller efter träning. Om du är uttömd natrium kommer din pump från träning nästan att vara obefintlig och natrium är nödvändigt för upptag av glutamin.
För att eliminera gissningar är Surge® Workout Fuel och Plazma ™ utformade med ideala elektrolytförhållanden för att stödja cellvolym och proteinsyntes.
Glutaminupptag i cellen orsakar cellvolym, vilket gör att muskelceller grundas för proteinsyntes. Som nämnts är en hel / volymiserad muskel en anabol muskel. Tillsammans med att driva aminosyratransport ökar cellvolymen också glykogensyntesen och hämmar proteinnedbrytning.(4-6)
Proteinsyntes undertrycks av utarmning av glutamin, vilket har enorma konsekvenser för idrottare med hård träning. Efter en intensiv träning är ett inflammatoriskt svar monterat, vilket gör att immunceller kan tränga in i trasig muskelvävnad för att börja reparera / bygga om.(14)
Glutamin tas så snabbt upp av immunceller att det anses vara ”immunförsvarets bränsle.”(15) Inte överraskande har det visat sig att intensiv träning orsakar plasmaglutaminutarmning.(16-18)
Av denna anledning ökar kraven på glutamin under perioden efter träning, där det lokala immunsvaret kan konkurrera om tillgängligheten av glutamin till primära muskelceller för aminosyraupptag och proteinsyntes.
Förladdning celler med glutamin kan också potentiellt minska "lag-time" associerad med leucinaktivering av proteinsyntes. Om du inte redan gör detta, ta 10-15 g glutamin eller glutaminpeptider omedelbart efter träningen. Eftersom BCAA är ett annat favoritsubstrat för muskelglutaminsyntes och har visat sig öka muskelglutaminproduktionen (19-21), är BCAA och leucin också användbara under perioden före träningen för att maximera endogen glutaminproduktion.
Redaktörens anmärkning: Hela Plamza / MAG-10-protokollet, med dess träning, träning och efter träning, uppfyller författarens rekommendationer för att ladda musklerna med BCAA och leucin.
Nyligen upptäcktes att konsumtionen av EAA ökar uttrycket för både System A och System L-aminosyratransportörer.(9) Viktigt är att detta händer på "post-transkriptionsnivå", vilket betyder på nivån för proteinsyntes, där befintliga mRNAs översätts till proteiner.
Kontrastera detta med “denovo”Proteinuttryck - där det kan ta 16 eller fler timmar att syntetisera, bearbeta och transportera nya mRNA - efter transkriptionell aktivering av proteinsyntes kan inträffa på några minuter till timmar, vilket gör att celler snabbt kan öka nivån på vissa proteiner vid behov.
Plötsligt har vi mer incitament att ha en gedigen näringsplan för peri-träning - EAA-intag under perioderna före och peri-träningen lönar sig stora efter träning genom att öka uttrycket för aminosyratransportörer, vilket gör att cellen får maximal aminosyra och aktivering av proteinsyntes.
Snabbabsorberande proteinisolat eller hydrolysat som Mag-10® Protein Pulsing Protocol ™ eller Plazma ™ under perioderna före och per träning är idealiska här.
Insulin är det mest anabola hormonet i kroppen. Tillsammans med direkt aktivering av proteinsyntes ökar insulin också translokation av System A-aminosyratransportörer till cellmembranet.(22)
Detta innebär att insulin får fler system A-transportörer att visas på cellmembranet, redo att driva mer glutamin in i cellen. Mer glutamin leder till mer cellvolym, vilket driver mer leucin in i cellen, vilket i slutändan leder till mer proteinsyntes.
Medan EAA ökar uttryck av AA-transportörer är det insulinsignalen som gör att de kan visas på cellytan, redo att skicka nya aminosyror in i cellen.
Detta är ännu en anledning till att kolhydrater före och peri är en bra idé om du inte är i extremt fettförlustläge: insulin ökar kapaciteten för cellulär aminosyratransport.
Kolhydrater ökar insulinnivåerna, men vissa aminosyror kan också användas för att förstärka insulinfrisättningen. Glutamin är en kraftfull aktivator av "inkretin" -hormoner som gör insulinproducerande celler i bukspottkörteln känsligare för glukos.(23) Glycin förstärker också insulinfrisättningen genom en annan mekanism.
Även om kolhydrater efter träning ensamt kommer att öka insulinnivåerna, kommer kombinationen av dessa insulinpotenserande aminosyror med kolhydrater att överbelasta bukspottkörteln för ännu mer insulinfrisättning. Även om det är bra att hålla insulinnivåerna på den nedre sidan för det mesta maximerar ökade insulinnivåer under peri-träningsperioden aminosyratransport, cellvolym och proteinsyntes samtidigt som proteinunderskottet undertrycks.
Den typ av intensiv anaerob träning som krävs för att bygga massor av kvalitetsmuskler leder till avsevärd laktatproduktion, vilket minskar muskel-pH. Detta leder till tidig muskeltrötthet och svaghet, men även vissa aminosyratransportörer, inklusive System A, hämmas av lågt pH.(13)
När muskel-pH är lågt reduceras upptagningen av aminosyror, vilket undertrycker mTOR-aktivering av proteinsyntes.(24) Det har också visats att låg hämning av aminosyratransportörerna av system A ökar proteinnedbrytningen.(25)
Det är här beta-alanin kommer in. Ökade muskelkarnosinnivåer fungerar som en naturlig syrabuffert, vilket förlänger den anaeroba tröskeln genom att begränsa minskningen av muskel-pH från träning.
Beta-alanin har en annan viktig funktion: att hjälpa till att upprätthålla proteinsyntesen och få det online snabbare efter intensiv träning genom att förhindra försvagning av aminosyratransport.
För att öka muskelkarnosinnivåerna, ta två tabletter Beta-7 ™ tre gånger om dagen.
Under intensiva träningspass minskar proteinsyntesen och proteinnedbrytning aktiveras. Detta är oundvikligt för alla hårda träningslyftare. Men i vilken utsträckning vi kan minimera de kataboliska effekterna av träning och ju snabbare vi kan återgå till ”anabola läget” under efterträningsperioden avgör i slutändan hur effektivt vi kommer att återhämta oss - och växa.
Tidpunkt för makronäringsämnen är viktigt, men det är ett sätt att nå målet. Cellvolym är den främsta drivkraften för aminosyratransport och proteinsyntes. Genom att förstå hur aminosyratransport sker och hur det regleras av cellvolym kan vi få mer leucin till trashed muskelceller snabbare, därigenom anstöt anabola eld och slutligen leda till bättre vinster.
Ovanstående strategier är effektiva, praktiska och baseras på den senaste vetenskapliga forskningen. Använd dem som en mall för att ta dina träningsförlopp till nästa nivå.
Jag ser fram emot dina frågor på LiveSpill!



Ingen har kommenterat den här artikeln än.